15510015856
细胞间通讯是指细胞之间相互通讯和传递各种信息的不同方式。这些交流方式包括许多不同的过程,而这些过程可以单独或同时发生,并且随着生理或病理环境的变化而改变。细胞间通讯的典型特征是释放可溶性因子,并影响邻近细胞的功能。然而,细胞之间还可以通过许多其他方式进行互相交流。在此,作者回顾了不同的细胞间通讯方式,包括在细胞衰老、机体衰老和衰老相关疾病中可溶性因子的作用。
要点
(1)细胞间通讯是生理和病理条件下的一个关键特征,目前假设几种细胞间通讯手段可同时发生或连续发生;
(2)通过可溶性因子进行细胞间通讯是目前研究最多的方式,然而重要的替代方式也逐渐被发现;
(3)衰老细胞会主动通过各种细胞间通讯手段与邻近细胞沟通,包括但不限于衰老相关分泌表型(SASP);
(4)大多数关于阻止可溶性因子释放的药理学药物的研究都忽视了这些药物对其他细胞通讯方式的影响。
词汇表
(1)Ferroptosis:铁死亡,指铁在细胞中积累引起的细胞死亡。
(2)Inflammaging:炎性衰老,指以慢性、低度炎症为特征的衰老。
(3)Inflammasome:炎症小体,指能够激活caspase-1并诱导先天炎症反应的免疫系统受体的聚集体。
(4)Senescence:衰老,指以一种或几种生物标志物(包括稳定的细胞周期阻滞和主动分泌体)为特征的表型的激活。
(5)Senescence-associated secretory phenotype (SASP):衰老相关分泌表型,指衰老细胞分泌的可溶性因子、生长因子和基质重塑酶。
(6)Senolytic drugs:延缓衰老药物,靶向杀死衰老细胞的药物。
(7)Senomorphics:抑制SASP的化学物质或药物。
(8)Syncytiotrophoblast:合胞体滋养层,滋养层细胞融合形成的胎盘外层。
(9)Trans-stable-isotope labelling of amino acids in cell culture (SILAC):细胞培养中氨基酸的跨稳态同位素标记,利用SILAC在细胞间转移蛋白质。
正文
一、细胞衰老:一个复杂和异质的表型
20世纪60年代,Moorehead和Hayflflick首次鉴定了一种称为衰老的细胞表型。在体外培养从人类供体中分离出的原代成纤维细胞时,他们观察到这些细胞会出现增殖停滞现象,并将这种表型称为细胞衰老。他们假设这种表型可以模拟衰老,并将其称为“培养皿中的衰老”。在体外发现了一种诱导提前衰老的现象,称为癌基因诱导的衰老,后来证实这种现象在体内抑制肿瘤进展方面发挥了作用。然而,直到2011年,Van
Deusen实验室才在细胞衰老和个体衰老激活之间建立了因果关系。作者在BubR1(有丝分裂检查点蛋白)缺失的早衰小鼠模型中发现某些器官中p16Ink4a的积累触发了早衰中常见的自然特征。有趣的是,他们首次发现p16Ink4a基因失活可以改善衰老。现在已经证实,细胞衰老是机体衰老的一个标志。此外,细胞衰老不仅是机体衰老的驱动因素,也是某些年龄相关疾病的驱动因素,如癌症、骨关节炎、动脉粥样硬化、阿尔茨海默病、慢性阻塞性肺病(COPD)和特发性肺纤维化(IPF)等。
虽然衰老的主要特征是细胞周期抑制剂p16INK4A和p21CIP的表达导致稳定的细胞周期阻滞,但细胞衰老对组织稳态的影响源于其高度主动的分泌特征。衰老相关分泌表型(SASP)被认为是细胞衰老的“灵魂”,因为它是高度主动的过程,并随着时间推移而改变其组分。它将产生有利的或不利的影响,而这取决于引发衰老的触发因素和背景。然而,由于在非常具体的情况下只确定了一些因素,SASP的特征还没有很好地被描述出来。还需要注意,衰老是一个复杂的异质细胞表型,能够在许多不同的环境中影响组织稳态,需要谨慎的将它标准化到某些标记(Box
1)。
Box 1
体外鉴定衰老细胞的指南
细胞衰老可由多种触发因素引起,包括端粒缩短、致癌应激、ROS和DNA损伤。进入衰老进程的原代细胞的主要表现是通过表达细胞周期抑制剂CDKN2A、CDKN2B和/或CDKN1A(分别编码p16INK4A、p15INK4B和/或p21CIP蛋白)诱导稳定的细胞周期阻滞,并缺乏增殖相关标记,如Ki67、BrdU或EdU(图1)。然而,由于这些标记不仅限于衰老细胞,也存在于未分化的体细胞中,因此应使用额外的标记来确认衰老表型。推荐鉴定三种以上的生物标志物,以证实衰老的激活。
另一个用于识别衰老的标记是衰老相关的β-半乳糖苷酶(SA-β-Gal)活性。SA-β-Gal的增加是由于衰老细胞中较高的溶酶体活性,这可能是由于衰老中溶酶体数量的增加或溶酶体活性的增加,并且可以通过特定的染色来检测。最后,还应检测其他标记,如DNA损伤、SASP的特定分泌体的释放或特定信号通路的激活,但这些标记更具争议,因为一些触发因素能够同时诱导这些标记的出现,如癌基因通过诱导H-RasG12V表达而诱导衰老。相反,其他触发因素,如发育衰老或αvβ3增加,不会引起一直被认为是衰老关键生物标志物的DNA损伤反应。这证实了并非所有的生物标志物都会在诱导衰老时同时表达,增加了衰老鉴定的复杂性。
需要注意的是,衰老是一种极其复杂和异质的细胞表型,在许多不同的背景环境中影响组织的稳态,标准化标记的推广可能是不利的,因为它很可能存在着与背景相关的、尚未被揭示的衰老特征。
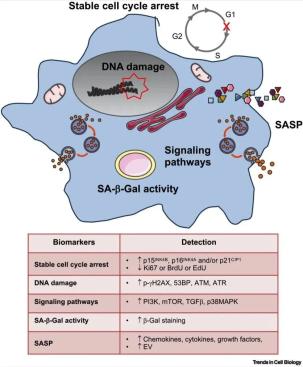 图1 衰老的生物学标记物。(EV,胞外囊泡;mTOR,哺乳动物雷帕霉素靶蛋白;SA-β-Gal,衰老相关β-半乳糖苷酶;SASP,衰老相关分泌表型;TGF-β,转化生长因子β。)
图1 衰老的生物学标记物。(EV,胞外囊泡;mTOR,哺乳动物雷帕霉素靶蛋白;SA-β-Gal,衰老相关β-半乳糖苷酶;SASP,衰老相关分泌表型;TGF-β,转化生长因子β。)
目前有两种主要的治疗策略来处理机体衰老和年龄相关疾病中的衰老细胞。第一个是通过选择性杀死衰老细胞(使用称为“senolytics”的药理学化合物)。第二个是通过使用senomorphics药物来消除细胞间通讯的有害影响,特别是SASP。在此,本文作者回顾了衰老细胞的不同细胞间通讯方式及其与机体衰老和一些年龄相关疾病的关系,将它们分为经典的、新发现的和非经典的细胞间通讯方式,包括可溶性因子和细胞外基质(ECM)重塑蛋白,也描述了细胞通讯的其他方式。文章重点关注了在不同的生物学背景和年龄相关的病理背景下,细胞间通讯是如何调节的及其功能。
二、SASP:经典和非经典细胞间通讯
SASP是一种衰老细胞特有的细胞间通讯手段。经典的SASP以衰老细胞分泌可溶性因子、生长因子和ECM重构酶为特征。然而,新发现的SASP和其他被称为非经典的细胞间通讯手段也在机体衰老和细胞衰老中被描述。在这里,作者回顾了经典、新发现的和非经典SASP的现有知识。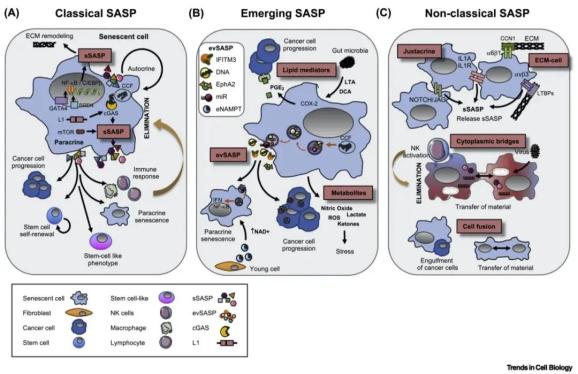 图2
细胞衰老和机体衰老过程中的细胞间通讯。(A)可溶性因子、生长因子和基质重构酶在经典可溶性衰老相关分泌表型(sSASP)模型中被释放。然而,sSASP需要受到严格的调控。在转录水平:BRD4和GATA4调控SASP的两个主要调控因子:NF-κB和C/EBPβ。sSASP也可以被LINE-1(L1)的逆转录转座元件驱动,这些逆转录转座元件在衰老过程中解脱抑制,而细胞质染色质片段(CCFs)通过活化环状GMP-AMP合成酶(cGAS)来驱动。哺乳动物雷帕霉素靶蛋白(mTOR)和下游信号通路也对该sSASP进行转录后调控。分泌的sSASP在微环境中起着多种作用,一方面,它改变了细胞外基质(ECM),可以通过自分泌信号增强衰老。另一方面,通过旁分泌信号也发挥许多功能。它可以促进癌症进展、干细胞自我更新和受损细胞中的干细胞样特征。同时还具有将衰老传递给邻近细胞的能力。重要的是,sSASP可以刺激先天和适应性免疫反应,有利于衰老细胞的消除。(B)新发现的SASP因子包括胞外囊泡(EVs)(evSASP)。由于胞外囊泡在衰老过程中包含物的高度异质性和多样性,新的功能不断涌现。一些因子逐渐被重视,如干扰素(IFN)和ephrin相关的抗氧化蛋白。此外,miRNAs(miRs)和CCF
DNA片段可以装载到evSASPs中。evSASPs还发挥旁分泌功能,促进癌症进展和诱导旁分泌引起的衰老。此外,已经发现代谢产物和脂质介质可以诱导氧化微环境或促进肿瘤进展。肠道微生物释放脂磷胆酸(LTA)和脱氧胆酸(DCA),它们通过COX-2和前列腺素E2
(PGE2)的释放驱动sSASP反应。(C)非经典SASP是指不受释放因子驱动的衰老过程中的细胞间通讯。细胞间通讯可以通过受体相互作用来介导,如白细胞介素-1受体(IL-1R)和白细胞介素-1
(IL-1A)或NOTCH1/JAG相互作用,两者均可诱导sSASP。此外,cell-ECM相互作用已被描述在衰老过程中发生。例如,
在αvβ3诱发sSASP时整合素α6β1可以与CCN1
(cysteine-rich蛋白1)相互作用进而通过活性氧(ROS)来诱导衰老。另外,称为细胞质桥的细胞质延伸也发生在细胞衰老过程中。有趣的是,衰老细胞和自然杀伤(NK)细胞之间发生的细胞质桥促进了衰老细胞的清除。细胞的融合也可以诱导细胞衰老,使融合细胞之间的物质交换成为可能。此外,衰老细胞可以吞噬癌细胞作为能量来源,维持衰老细胞的高代谢需求。(eNAMPT,胞外烟酰胺磷酸核糖转移酶;EphA2,肝配蛋白A;;IFITM3,干扰素诱导的跨膜蛋白3;LTBP,潜活相关的TGF-β结合蛋白。)
图2
细胞衰老和机体衰老过程中的细胞间通讯。(A)可溶性因子、生长因子和基质重构酶在经典可溶性衰老相关分泌表型(sSASP)模型中被释放。然而,sSASP需要受到严格的调控。在转录水平:BRD4和GATA4调控SASP的两个主要调控因子:NF-κB和C/EBPβ。sSASP也可以被LINE-1(L1)的逆转录转座元件驱动,这些逆转录转座元件在衰老过程中解脱抑制,而细胞质染色质片段(CCFs)通过活化环状GMP-AMP合成酶(cGAS)来驱动。哺乳动物雷帕霉素靶蛋白(mTOR)和下游信号通路也对该sSASP进行转录后调控。分泌的sSASP在微环境中起着多种作用,一方面,它改变了细胞外基质(ECM),可以通过自分泌信号增强衰老。另一方面,通过旁分泌信号也发挥许多功能。它可以促进癌症进展、干细胞自我更新和受损细胞中的干细胞样特征。同时还具有将衰老传递给邻近细胞的能力。重要的是,sSASP可以刺激先天和适应性免疫反应,有利于衰老细胞的消除。(B)新发现的SASP因子包括胞外囊泡(EVs)(evSASP)。由于胞外囊泡在衰老过程中包含物的高度异质性和多样性,新的功能不断涌现。一些因子逐渐被重视,如干扰素(IFN)和ephrin相关的抗氧化蛋白。此外,miRNAs(miRs)和CCF
DNA片段可以装载到evSASPs中。evSASPs还发挥旁分泌功能,促进癌症进展和诱导旁分泌引起的衰老。此外,已经发现代谢产物和脂质介质可以诱导氧化微环境或促进肿瘤进展。肠道微生物释放脂磷胆酸(LTA)和脱氧胆酸(DCA),它们通过COX-2和前列腺素E2
(PGE2)的释放驱动sSASP反应。(C)非经典SASP是指不受释放因子驱动的衰老过程中的细胞间通讯。细胞间通讯可以通过受体相互作用来介导,如白细胞介素-1受体(IL-1R)和白细胞介素-1
(IL-1A)或NOTCH1/JAG相互作用,两者均可诱导sSASP。此外,cell-ECM相互作用已被描述在衰老过程中发生。例如,
在αvβ3诱发sSASP时整合素α6β1可以与CCN1
(cysteine-rich蛋白1)相互作用进而通过活性氧(ROS)来诱导衰老。另外,称为细胞质桥的细胞质延伸也发生在细胞衰老过程中。有趣的是,衰老细胞和自然杀伤(NK)细胞之间发生的细胞质桥促进了衰老细胞的清除。细胞的融合也可以诱导细胞衰老,使融合细胞之间的物质交换成为可能。此外,衰老细胞可以吞噬癌细胞作为能量来源,维持衰老细胞的高代谢需求。(eNAMPT,胞外烟酰胺磷酸核糖转移酶;EphA2,肝配蛋白A;;IFITM3,干扰素诱导的跨膜蛋白3;LTBP,潜活相关的TGF-β结合蛋白。)
(一)经典SASP:可溶性因子或sSASP
sSASP对组织稳态既有利又有害。因此,需要对sSASP进行严格调控。sSASP的主要驱动因素之一是持续的DNA损伤反应。两个主要调控SASP的因素是:NF-κB和C/EBPβ,而NF-κB受转录因子GATA4的调控(图2A)。然而,独立于p38
MAPK和胞质染色质片段(CCFs)的非典型DNA损伤反应也可以诱导sSASP。这些DNA片段可以在衰老过程中从细胞核中释放出来,激活干扰素基因(STING)通路的抗病毒环状GMP-AMP合酶(cGAS)刺激因子。不过,p38
MAPK和CCF激活sSASP需要通过NF-κB信号。
奇怪的是,炎症小体的激活也可以调控sSASP。这是由白介素(IL)-1的信号转导和表达介导的,主要参与旁分泌衰老信号。此外,IL-1A受到哺乳动物雷帕霉素(mTOR)抑制靶点的调控。mTOR选择性地通过IL-1A转录后调控多个sSASP因子,也通过蛋白合成因子4EBP1调控MK2[丝裂原活化蛋白激酶激活蛋白激酶2(MAPKAPK2)]来调控sSASP。
另外,表观遗传改变也可以调节sSASP。最近的一项研究发现,在细胞衰老和衰老过程中逆转录转座因子LINE-1(L1)的抑制。这进而激活了cGAS/STING,从而激活了sSASP。此外,sSASP还受到染色质重塑以及BRD4向sSASP基因附近超增强子的招募的调控。总的来说,由于功能异常的表达和释放可以导致病理状态,因此sSASP要受到严格的调控。
长期以来,人们认为sSASP的成分会随着时间的推移而改变。直到最近,一项全面的研究才发现sSASP包含两种不同的功能。第一是富含抗炎转化生长因子(TGF)-β的sSASP。这种特殊的sSASP由膜结合蛋白NOTCH1调控,NOTCH1的表达在衰老的初始阶段增加,并介导C/EBP抑制。第二是随着时间的推移,NOTCH1表达降低,导致C/EBP蛋白的活化,进而诱导促炎sSASP产生。




 大
大